[ 27: 腸管出血性大腸菌の病原性制御因子による遺伝子発現制御機構 ]
O-157 のゲノムはモザイク構造。
いろいろな外来性遺伝子がはいっているために場所によって GC 含量が違う、とか。
では、外来性遺伝子は病原性を発揮するためにどのように制御されているのか?
ふつうは大腸菌の H-NS によって外来性遺伝子が silencing されており、これを knockout すると O-157 でも病原性があがる。
Pch がある・ない条件で、H-NS がどこにくっついているかを調べた。
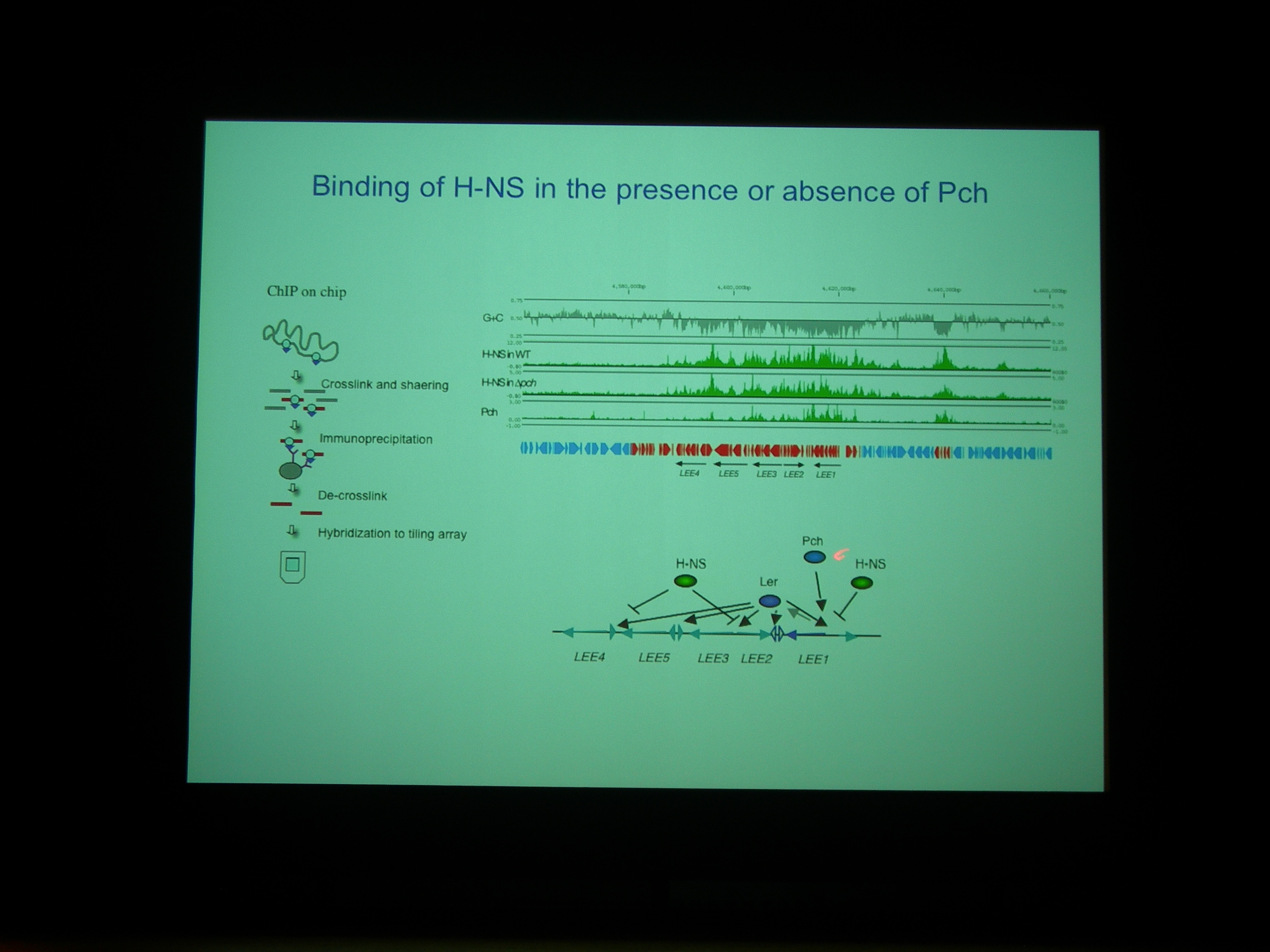
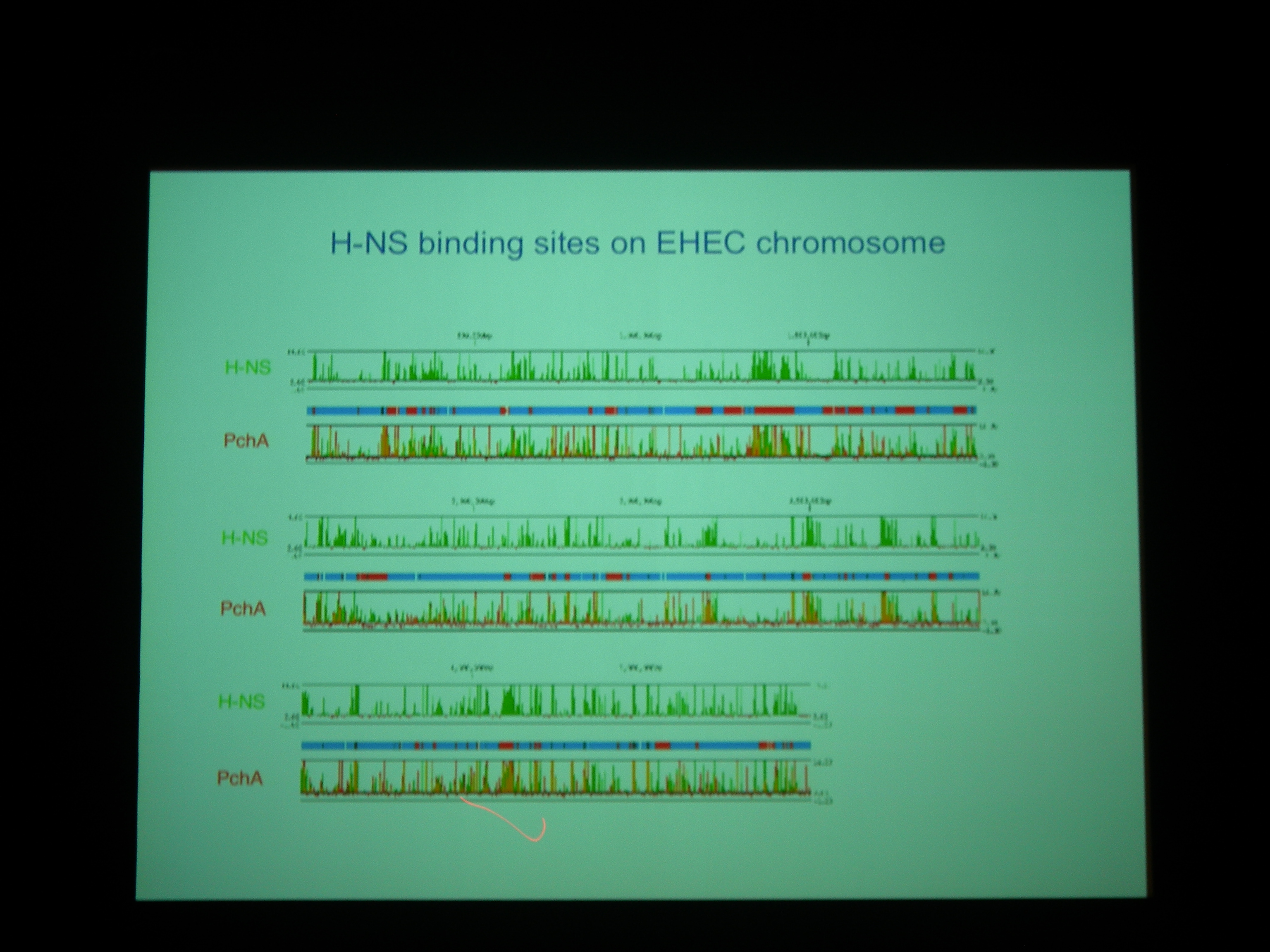
H-NS と Pch の binding site は非常によく似ている。Pch+ でも H-NS の bind する量はあまりかわらないが、よくみると変わっている (H-NS の bind が若干下がる) ところがあり、そこにはLEE1-5 などの遺伝子がコードされている。
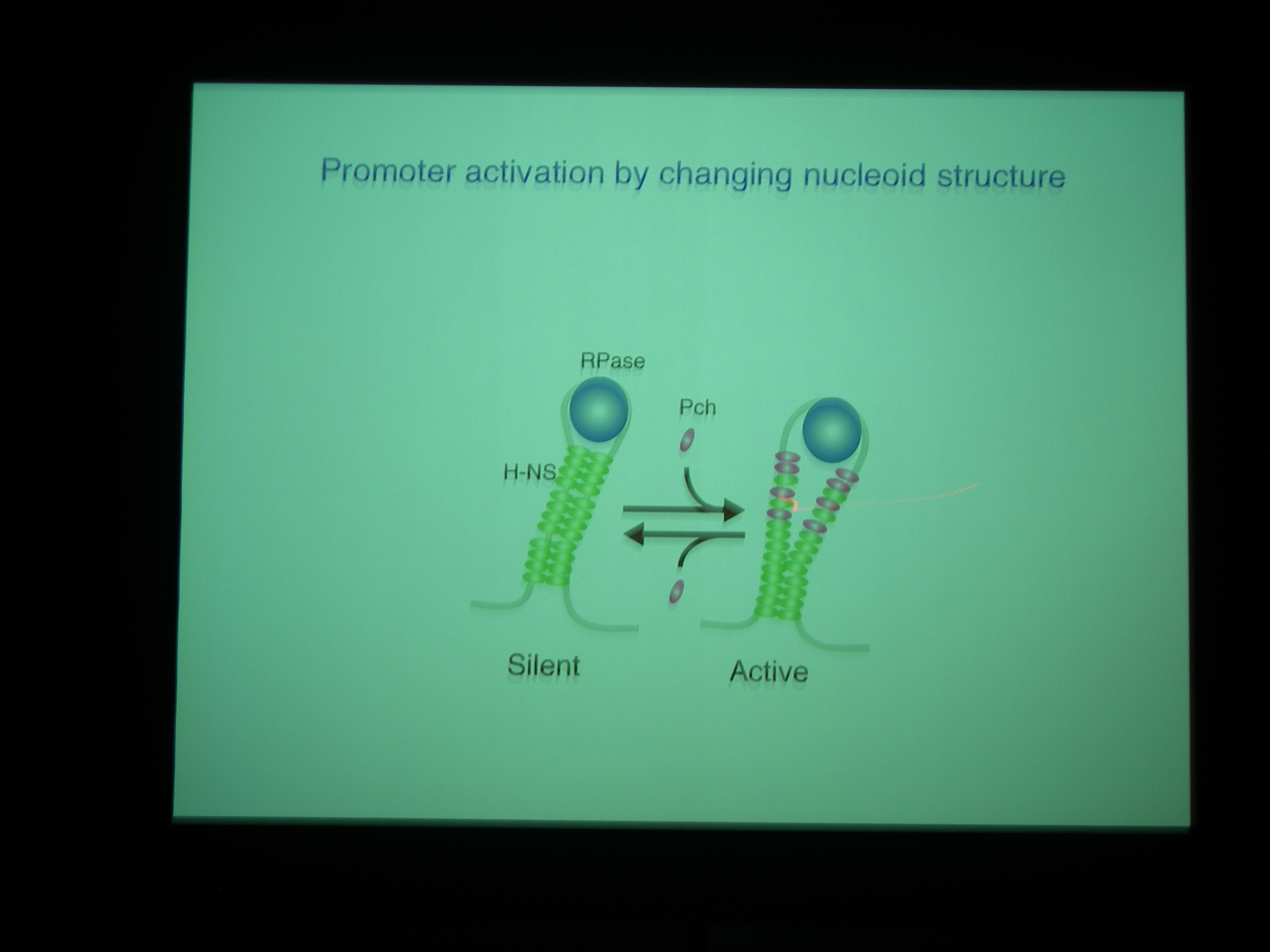
PchA の発現量を増やしてやると、LEE の発現量も増える→病原性上がる。
Pch がついているときは、H-NS が小さな固まりで bind している模様。H-NS がいっぱいくっついているときは転写を阻害しているが、Pch がはいってきちゃうと転写されるようになっているのではないか。
Pch は病原性遺伝子だけを制御している? → ほかのところにもつきます。あと、H-NS や Pch 以外のファクタもあると思う。
H-NS や Pch がつく場所のコンセンサス配列みたいなのはわかるの? → AT rich だということくらいしかいえない。タンパクのサイズは 10kd くらい。
[ 28: 枯草菌の複数シグマ因子による転写制御ネットワーク解明の試み ]
たくさんのシグマ因子がたくさんの遺伝子の調節をこまかくやっている。
ECF family が半分くらいあるといわれているが、機能が (環境応答らしい、ということくらいしか) よくわからない場合が多い。発現ストレスが似ていたり、プロモータ領域が似ていたりして、つぶして解析したりすることがむずかしい。
7つあるので、それをまず全部破壊する (抗生物質耐性の遺伝子とかがあるので、マーカーありでやるのは難しい)。
WT でも、胞子形成後に溶菌する。
ECF7 つと、sigI を壊すと溶菌する率があがることがわかった。
いくつかの ECF と sigI を破壊して、同じような様相を呈するケースを探している。
– 溶菌は2成分制御系 (YvrG/ YvrHb とかで、最終的にオートリシン LytC が発現)。
– で、yvrG は sigI に制御されているので、sigI を壊すと最終的に溶菌の制御も壊れる。
– yvr* を IPTG で転写誘導したり、lytC を壊すことで溶菌は抑圧できる。
ECF family は環境応答だけでなく恒常性維持とかにも関わっているらしい。
大腸菌の programmed cell death との関わりは? → やっぱりシグマ因子だけど、大腸菌ではシグマ因子の発現が細胞死を誘導するので、逆になっている。
[ 29: 枯草菌におけるフラボノイド応答性転写制御系の機能解析 ]
気絶。すみません…
[ 30: シアノバクテリアSynechococcus elongatus PCC 7942 における二成分制御系シグナル伝達ネットワークの解析 ]
多くの場合は2成分制御系のヒスチジンキナーゼとレスポンスレギュレータはゲノム上に並んで operon 構造を持っているが、シアノバクテリアでは並んでおらず、orfan になっている。
Operonになっているものもあるが、orphan なものですべてのシアノバクテリアに保存されているものが 7 つ。NblS, RpaB, Syc1065, Ycf29 はシアノバクテリアの必須遺伝子であり、紅藻からも同定されている重要な遺伝子。
相互作用を網羅的に調べてみた。16遺伝子間、10組のパートナーを同定。ヒスチジンキナーゼ同士の組もある。
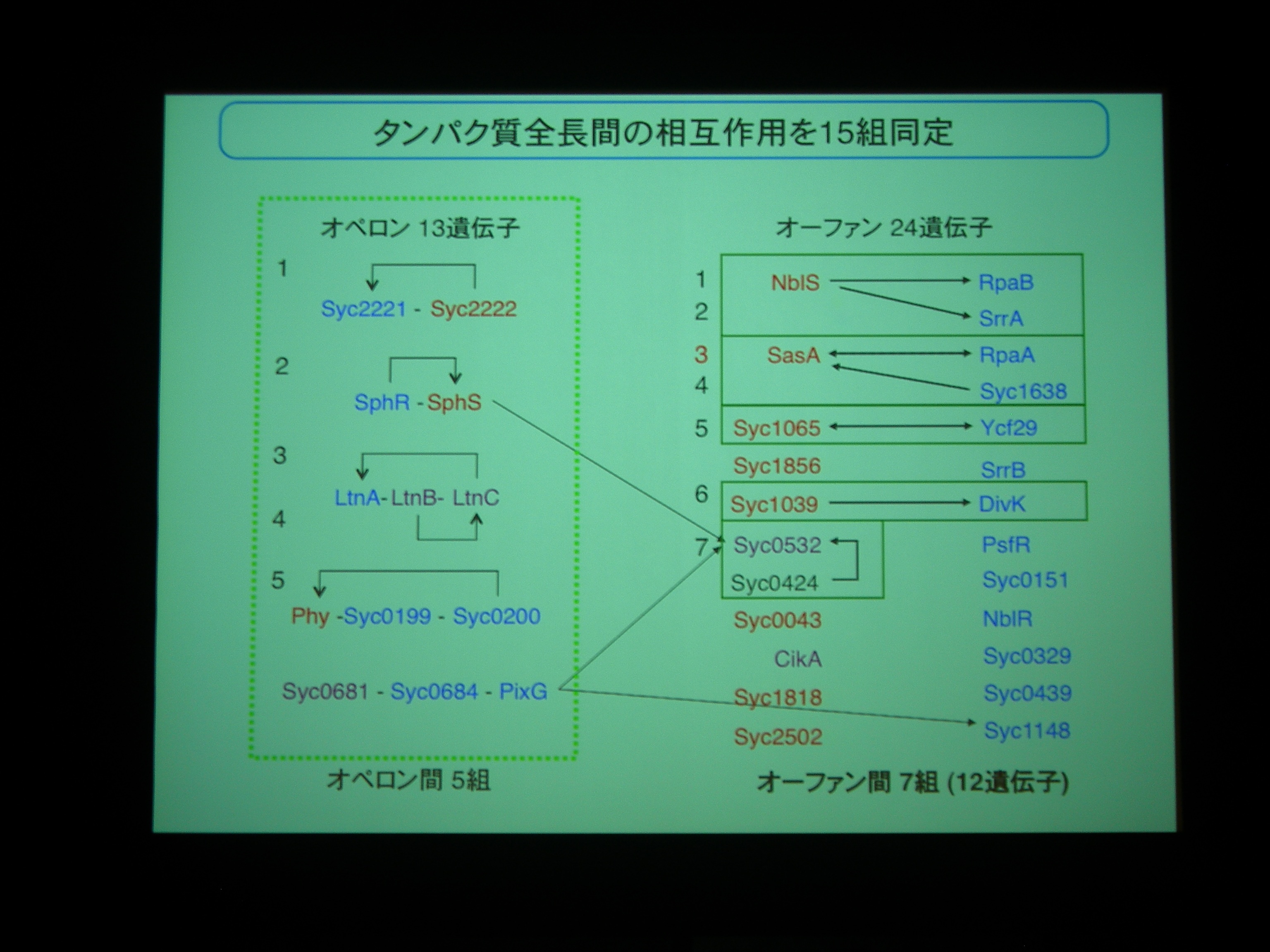
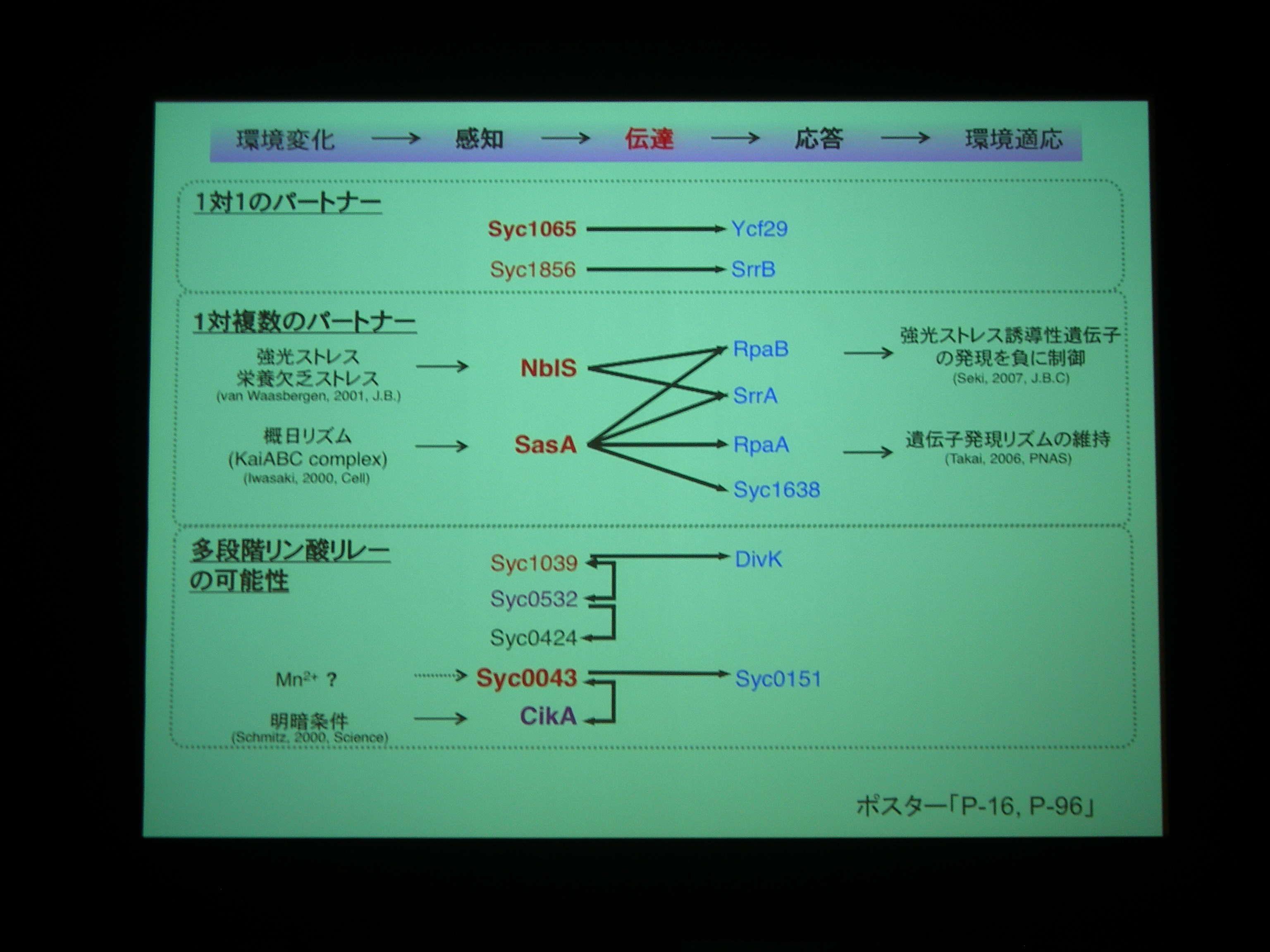
1対複数のパートナーとかを構成するためには operon より orfan でいたほうが都合がよいのかも? ということでした。
[ 31: Rhodobacter sphaeroidesの光合成から好気呼吸へのエネルギー代謝転換時における全ゲノム転写プロファイルの動的変化 ]
染色体2つとプラスミド5個。
嫌気性条件下で特異的に光合成 (自己栄養)。酸素があれば呼吸する。酸素のある・なしで定常条件になっている奴らは充分に研究されているが、transition state は?
Gene chip を使って、嫌気性条件から好気性条件へ遷移する transition state における発現をしらべた。NADH dehydrogenase なども嫌気性条件と好気性条件で別々の遺伝子を使ったりしているところがおもしろい。
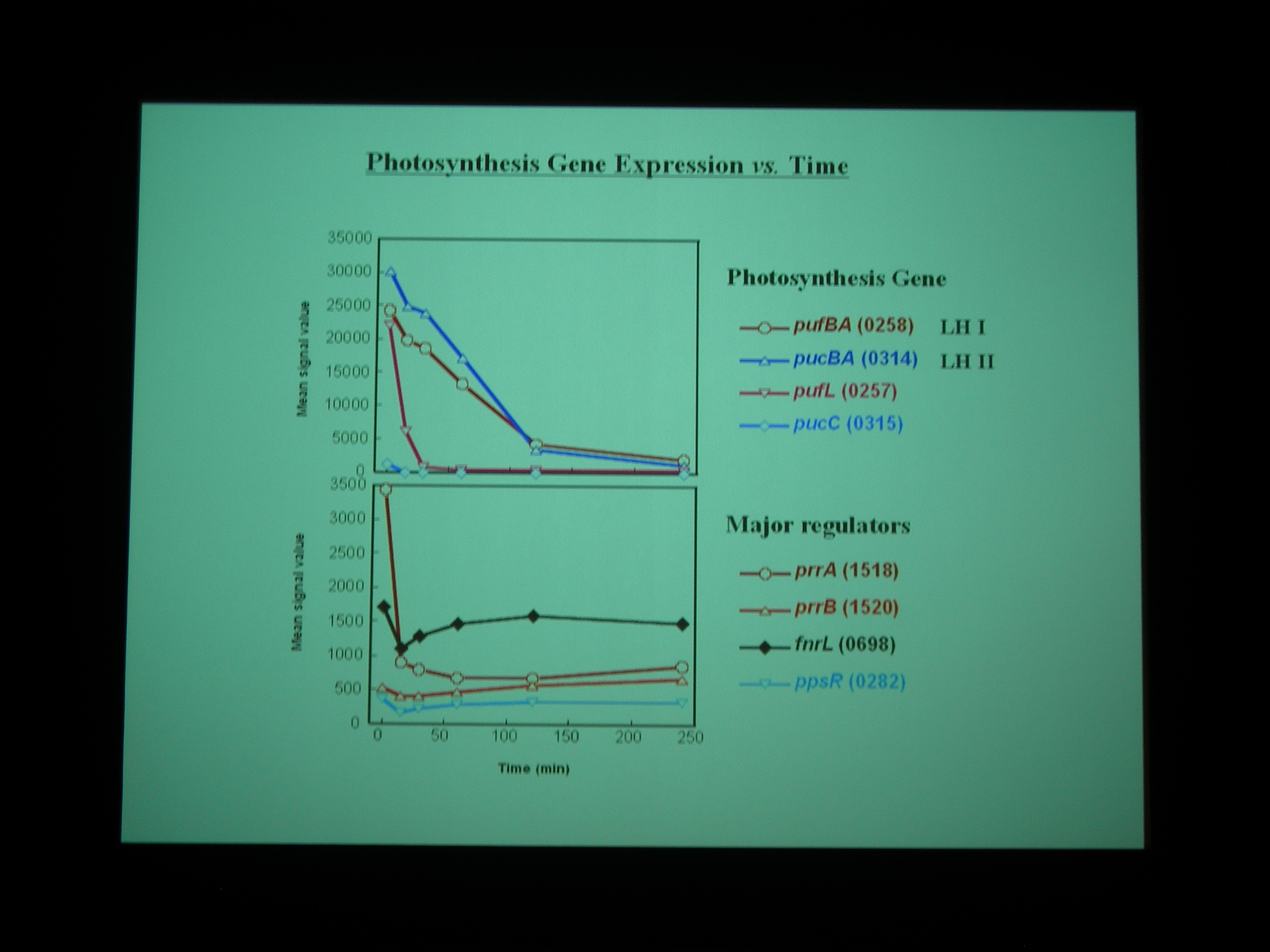
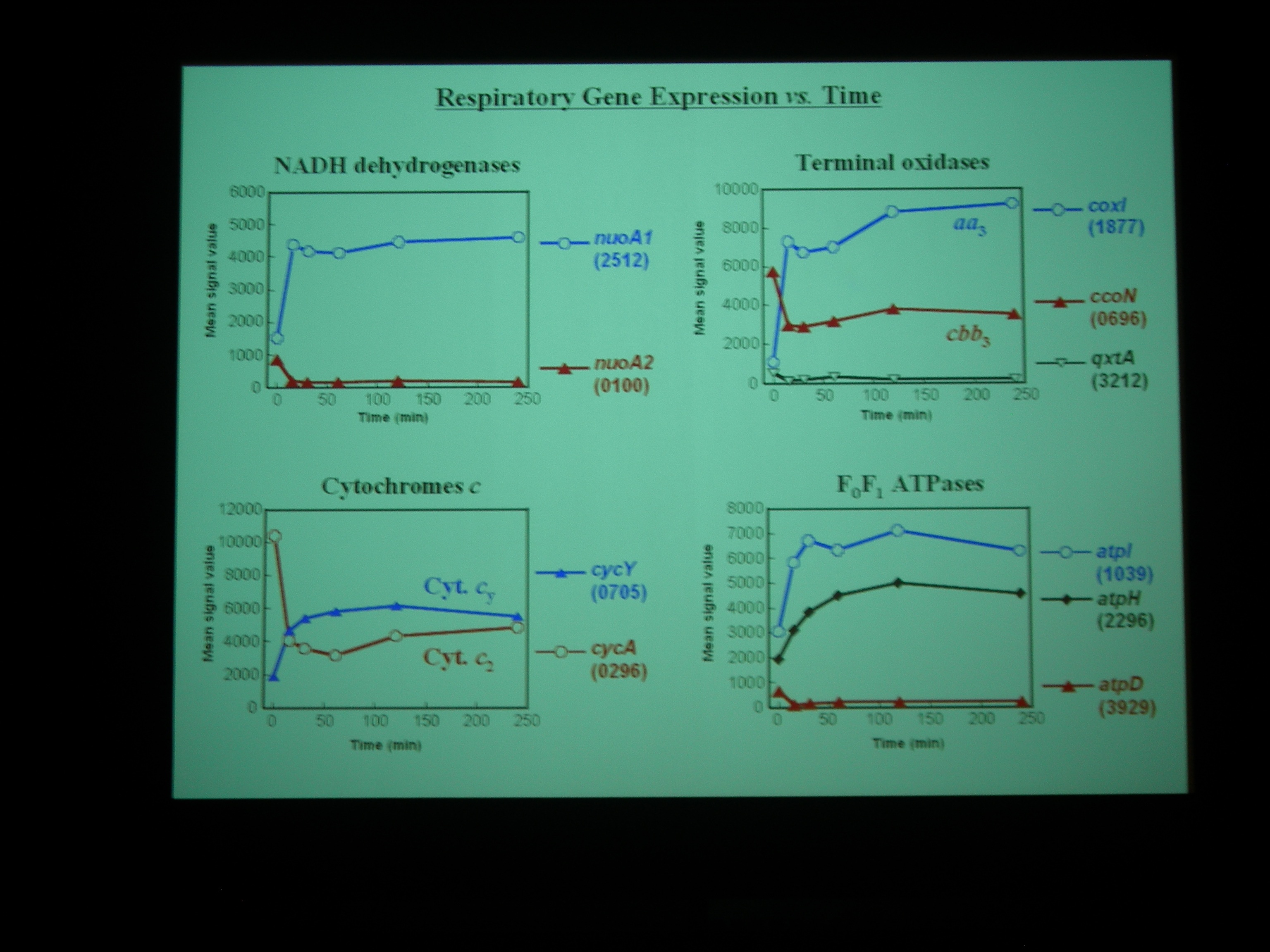
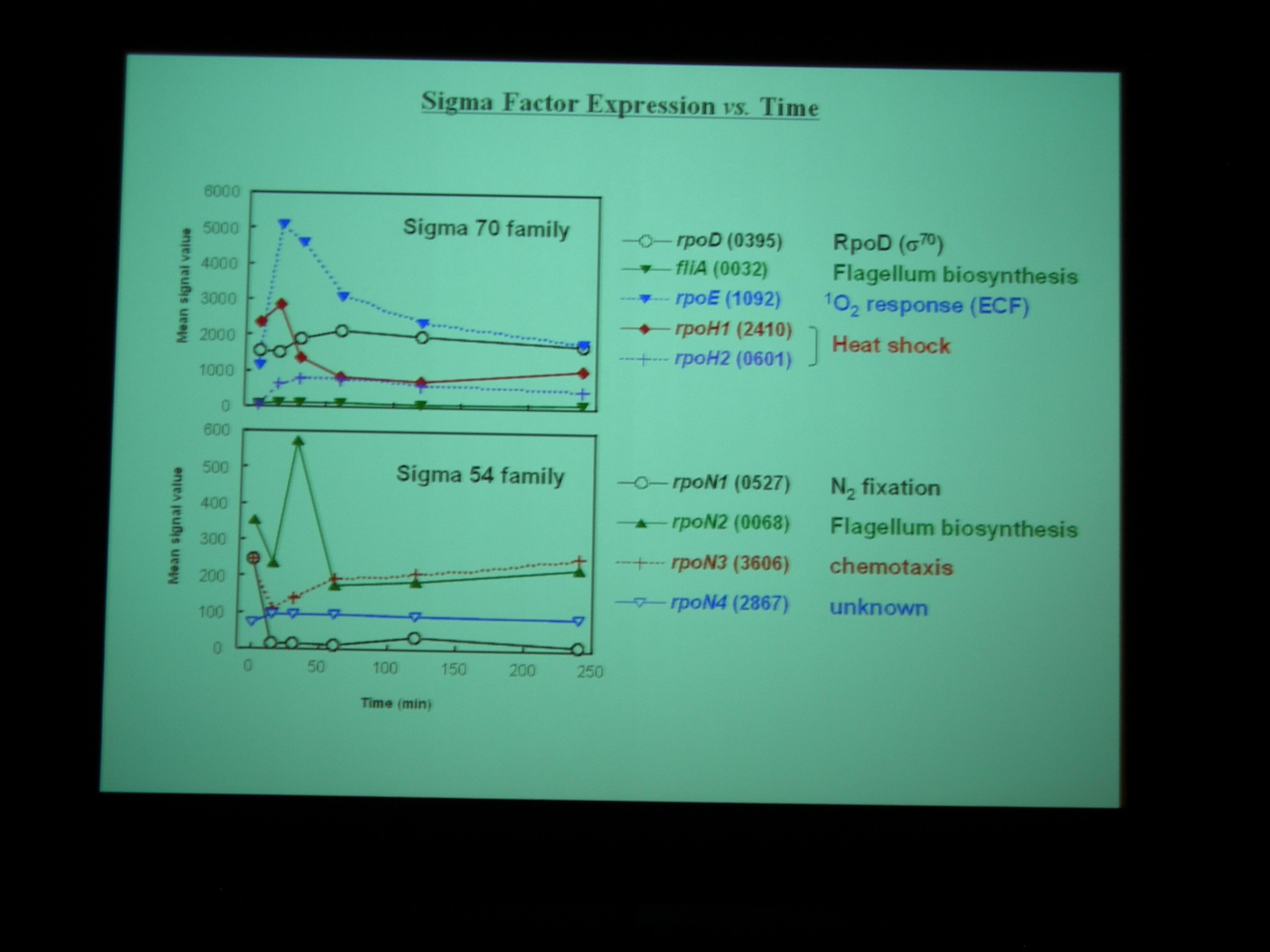
Class I – IV の遺伝子に発現をみていると、酸素を与えたところで細胞周期が同期している。
好気性条件になると、最初に光合成で作った糖を食べて、それから培地の炭素源を使う。

いろいろな生き物がいるねぇ・・・・
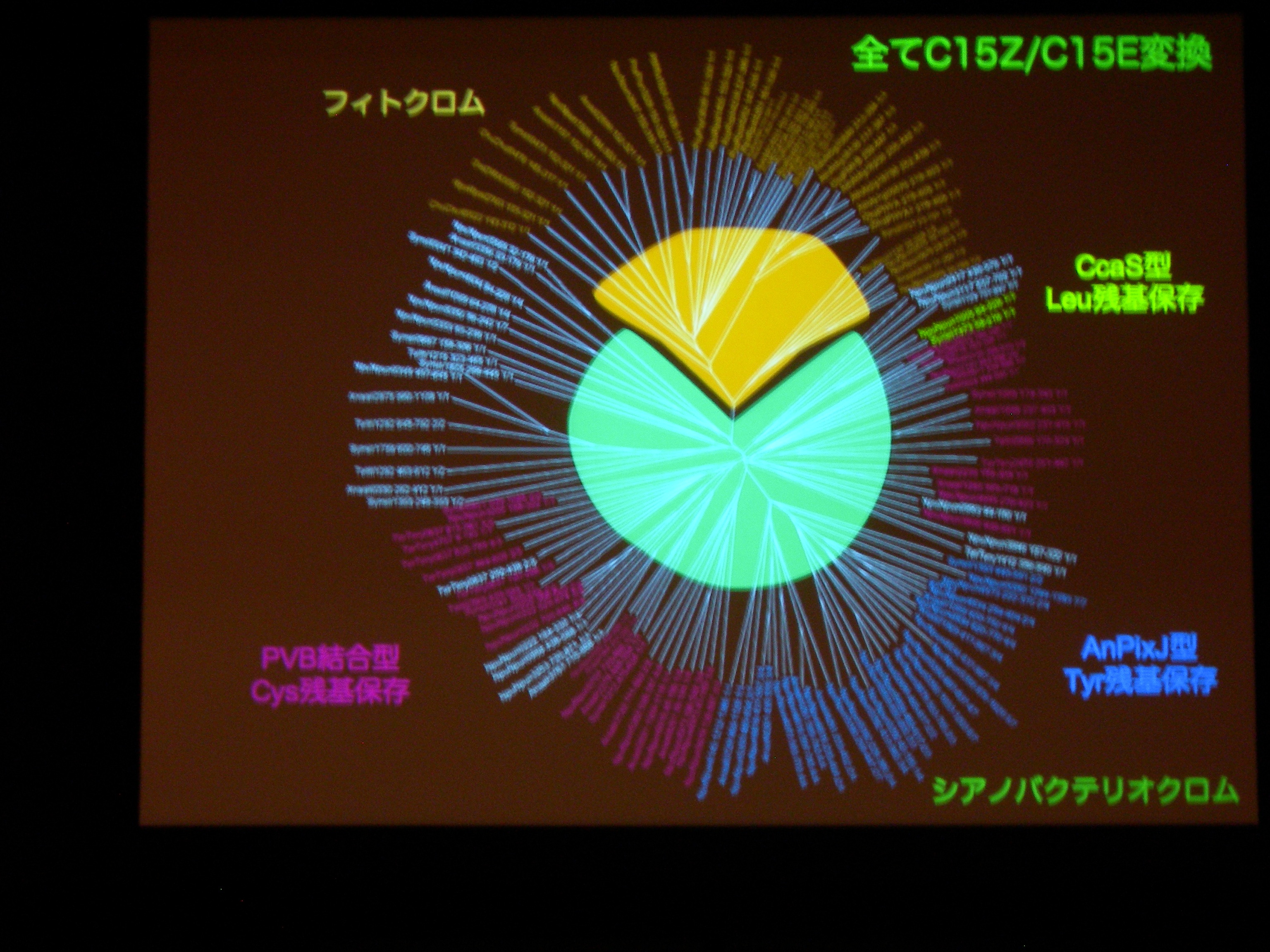
[ 32: 構造-機能解析によるシアノバクテリオクロムの多様性と普遍性の理解 ]
光受容体 (Phytochrom) に似た、シアノバクテリアにしかないシアノバクテリオクロムというのが注目されている。ものすごくたくさん種類があり、吸収スペクトルが違う。
AnPixJ の Pr 型を結晶化することに成功。フィトクロムの Pr 型と比べると、タンパク質と色素の位置関係が違うものの、タンパク質どうし、色素どうしはよく似ている。でも、色素のまわりの水素結合のようすはだいぶ違う。
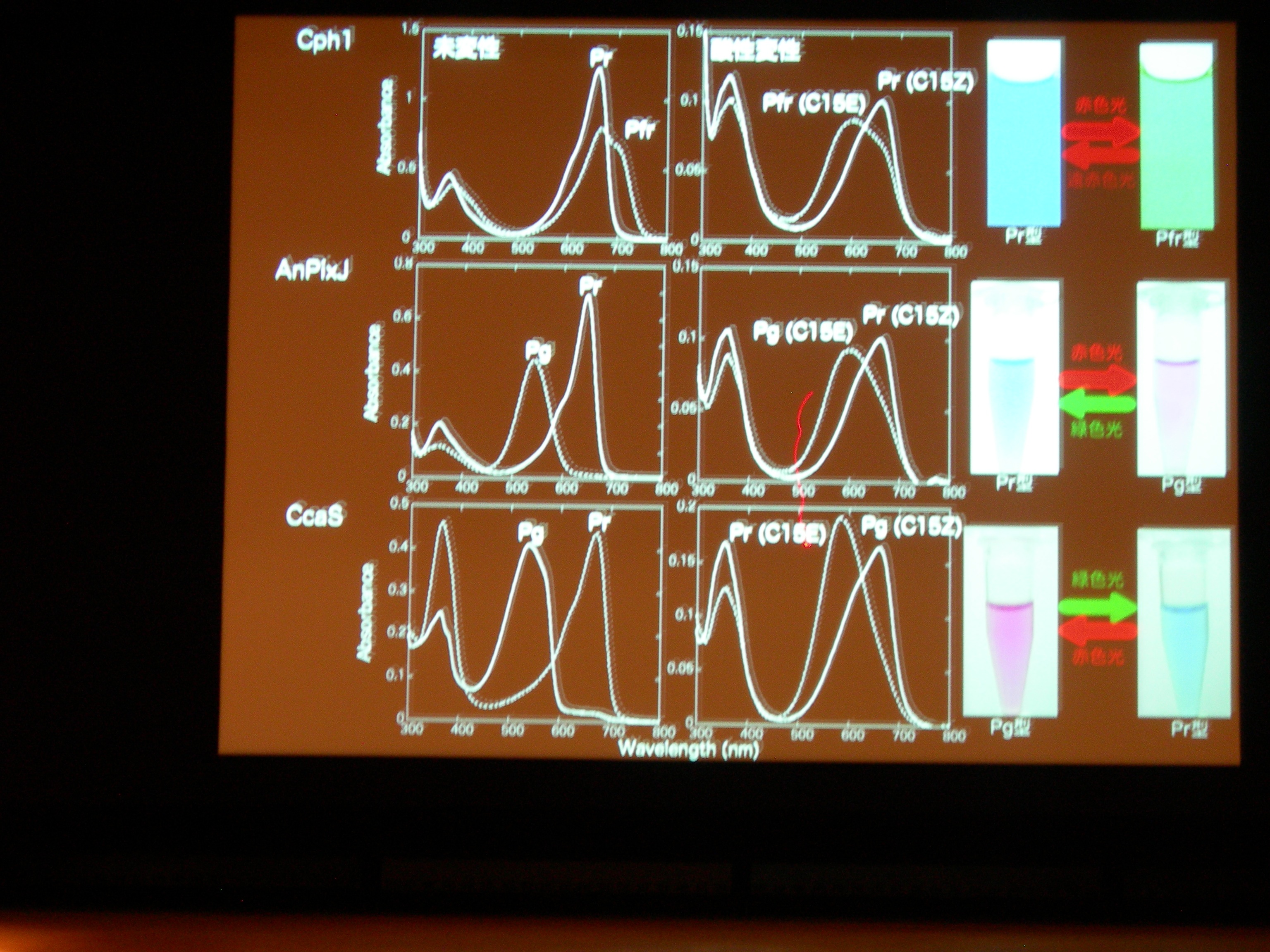
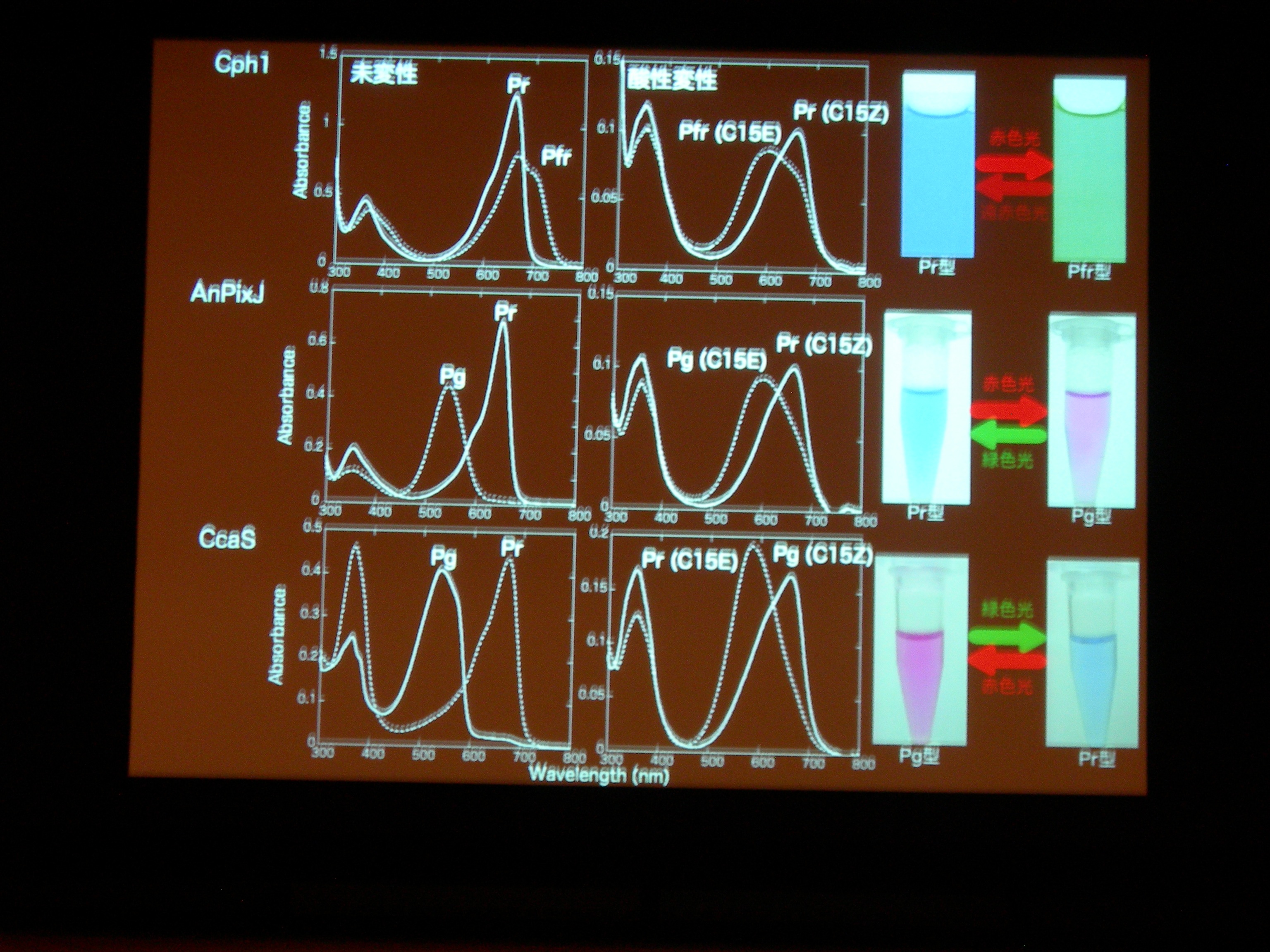
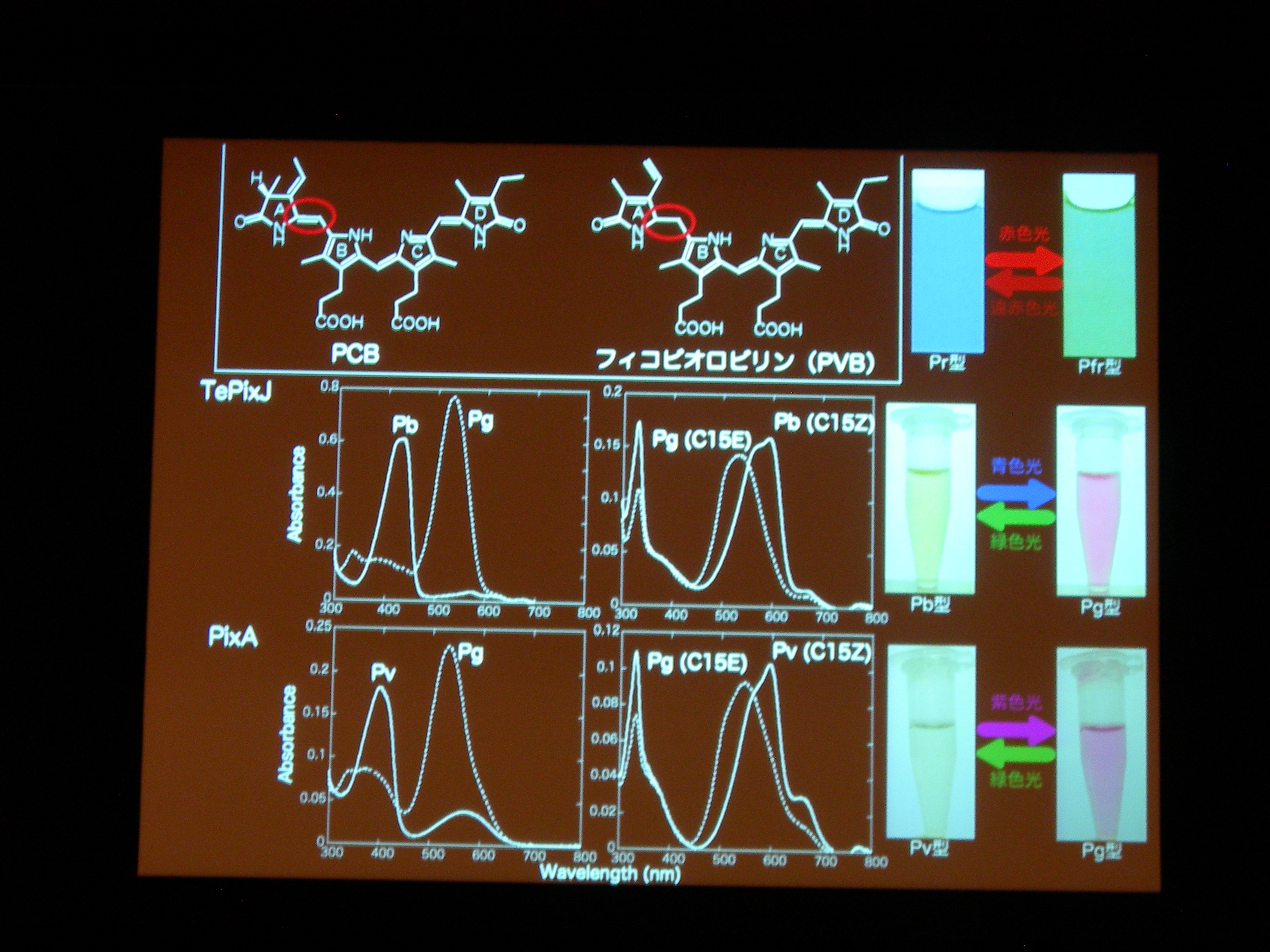
タンパク質と色素の共進化ですが、祖先型みたいなのはわかるかしら? → おもしろい suggestion ですが、とてもむずかしい。
ここまで多様な理由は? → シアノバクテリアは緑色光を吸収する必要があったりとか、そういうことが理由の一つとして考えられる。