[ 21: Replication dynamics of the Vibrio chromosomes affect gene dosage, expression and location ]
ふたつある染色体のそれぞれの複製のときにどんな遺伝子がたくさん発現してるか、とかを実験的に確認。
片方の染色体は重要で安定しており、もう片方は flexible.
小さい方の origin は plasmid かも、というお話。

[ 22: プラスミド分配の目的地を規定するモータータンパク質、ParA ]
細胞分裂のときにプラスミドはどうやって分配されるのか?
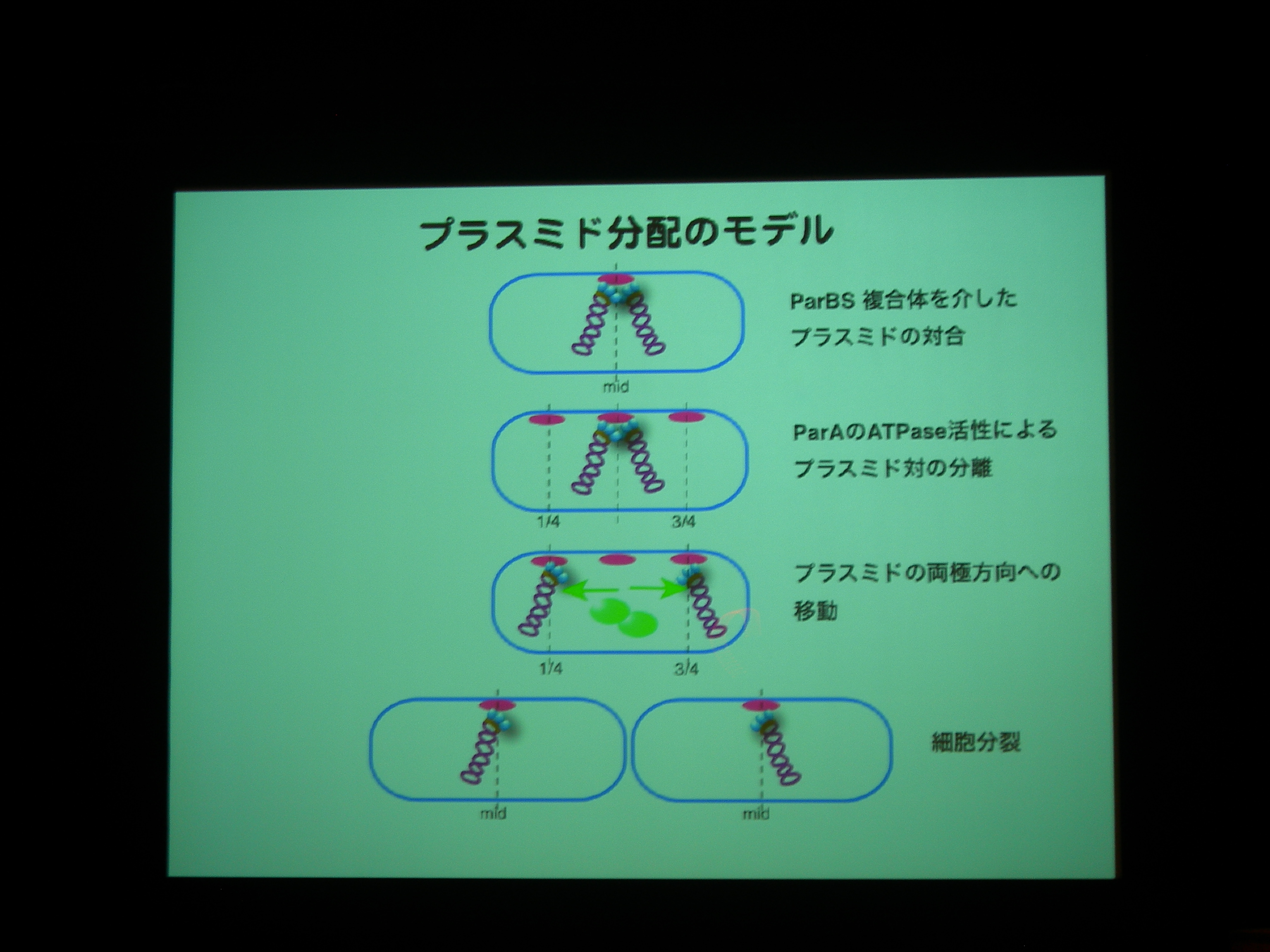
プラスミド分配に必要なタンパク質には actin type のものと walker type のものがある。
actin type はうにょーっと伸びるわけだが、walker type は細胞の極から極へ移動し、プラスミドを連れていくっぽい。
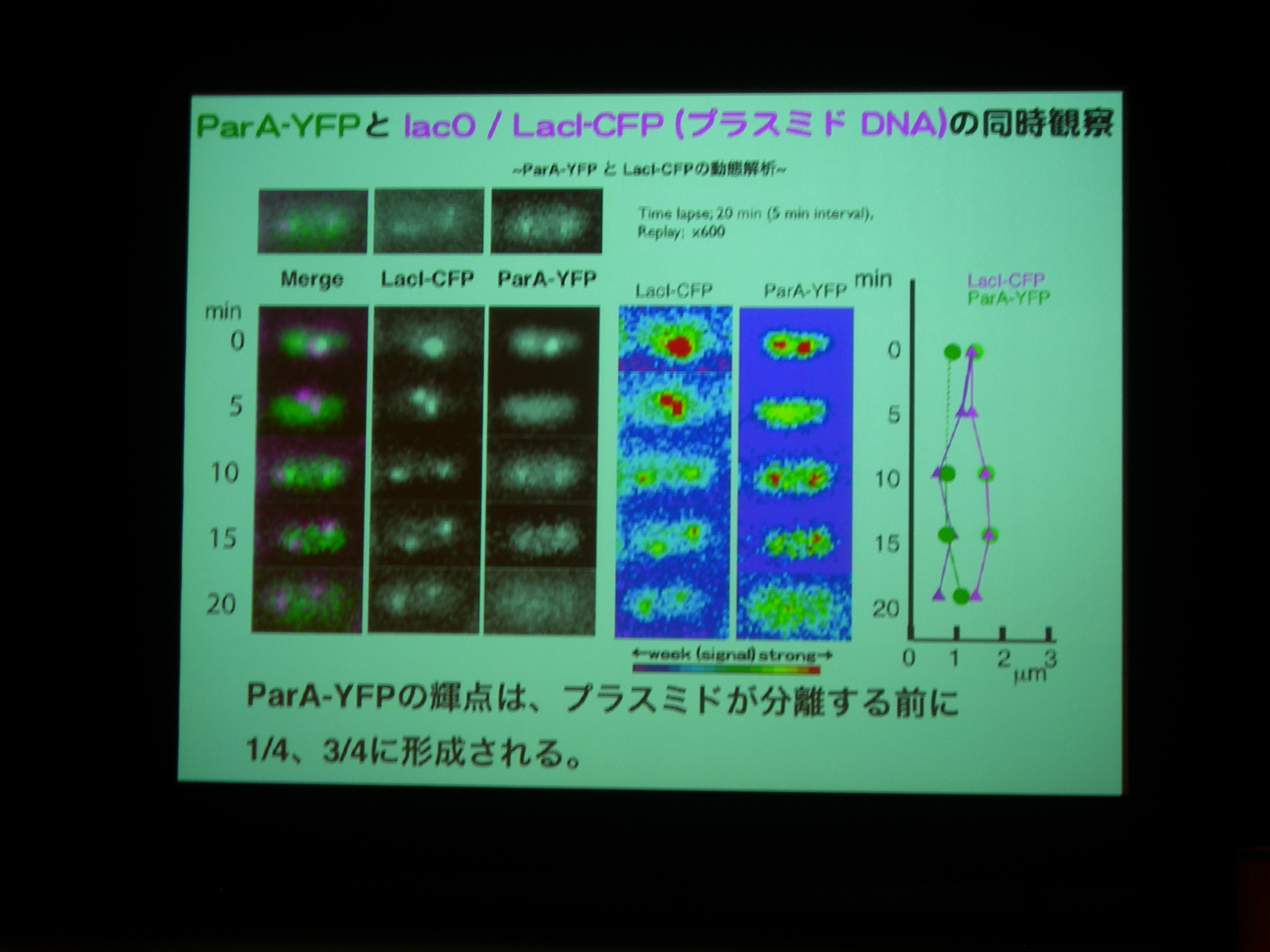
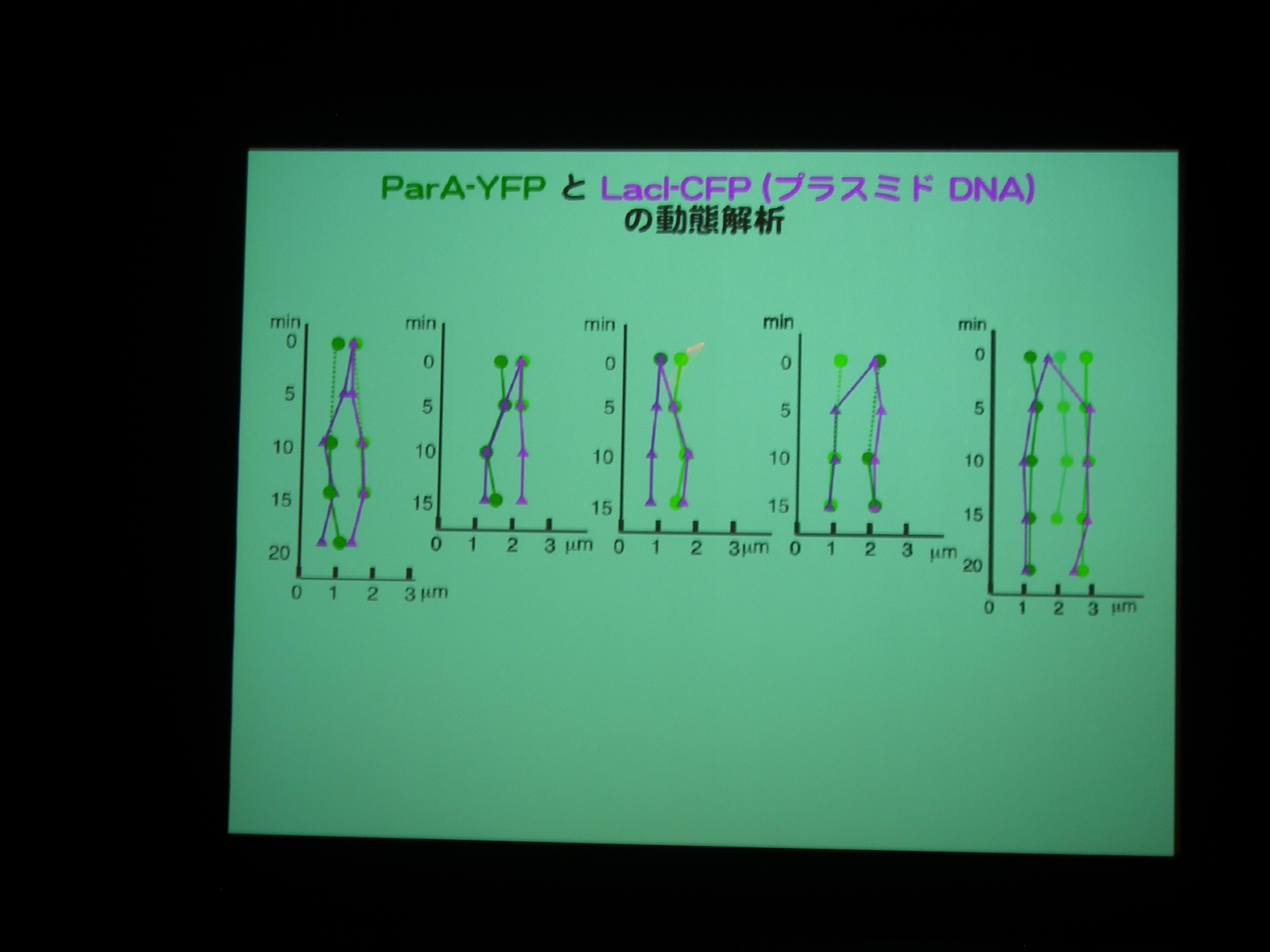
ParA protein に YFP, Plasmid に LacI promoter + CFP をつけて動態を観察。
ParA を追いかけると、細胞中央に輝点が現われ、1/4 地点に移動した後に 3/4 地点にも現れる。
プラスミドがふたつになる前に ParA がふたつになっている?


ParA を壊したり、ParA の ATPase を壊したりすると、局在しなくなる。
ParA を overexpression させたら? → 安定しなくなっちゃいます
[ 23: 枯草菌の50Sリボソームサブユニットの生合成におけるGTP結合タンパク質の結合モデルの提案 ]
50S subunit の後期生合成の関する研究。なんかすごい。
でも、完璧にウェットなお話でした。わからんです。
[ 24: Phylum Bacteroidetesに位置する細菌の菌体外タンパク分泌機構と滑走運動機構との関連性 ]
自分で糖発酵をせず、エネルギーを外界に依存。
表面に強力なプロテアーゼ (ジンジパイン: これ自体病原因子?) を持つ。たいていは膜結合な複合体になる。
分泌機構はよくわかっていないが、変異株にはジンジパインを外側に出すことができず、前駆体を細胞内に蓄積してしまうものがある。
P. gingivalis 輸送系に影響を与える変異を F. johnsoniae にいれてやると、滑走しなくなる。
膜タンパクであるところが共通だけど、それ以上のことについてはまだよくわからない。
[ 25: 分裂酵母Meu14は前胞子膜開口部を量依存的に制御する ]
分裂酵母の胞子形成。減数分裂後に核を包み込むように膜が生成される。
膜が袋状になるしかけが不思議。膜の先端には Meu14 というタンパク質があり、それが巾着の口をとじるところに関係している?
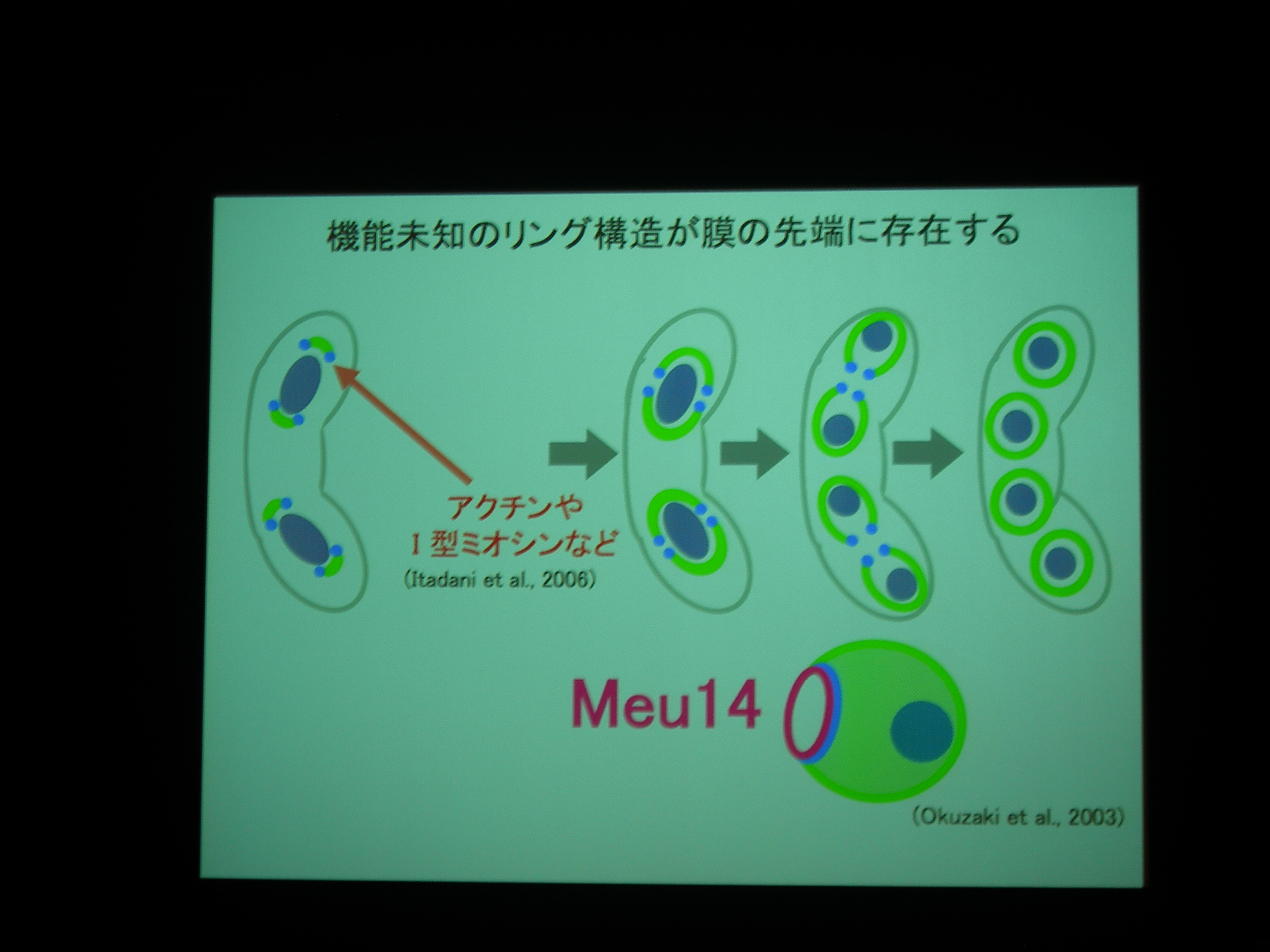
膜と核と Meu14 を蛍光で観察。
Meu14 がないと、開口が維持されず、さっさと口を閉じてしまって、核を包み込むことができなくなる。
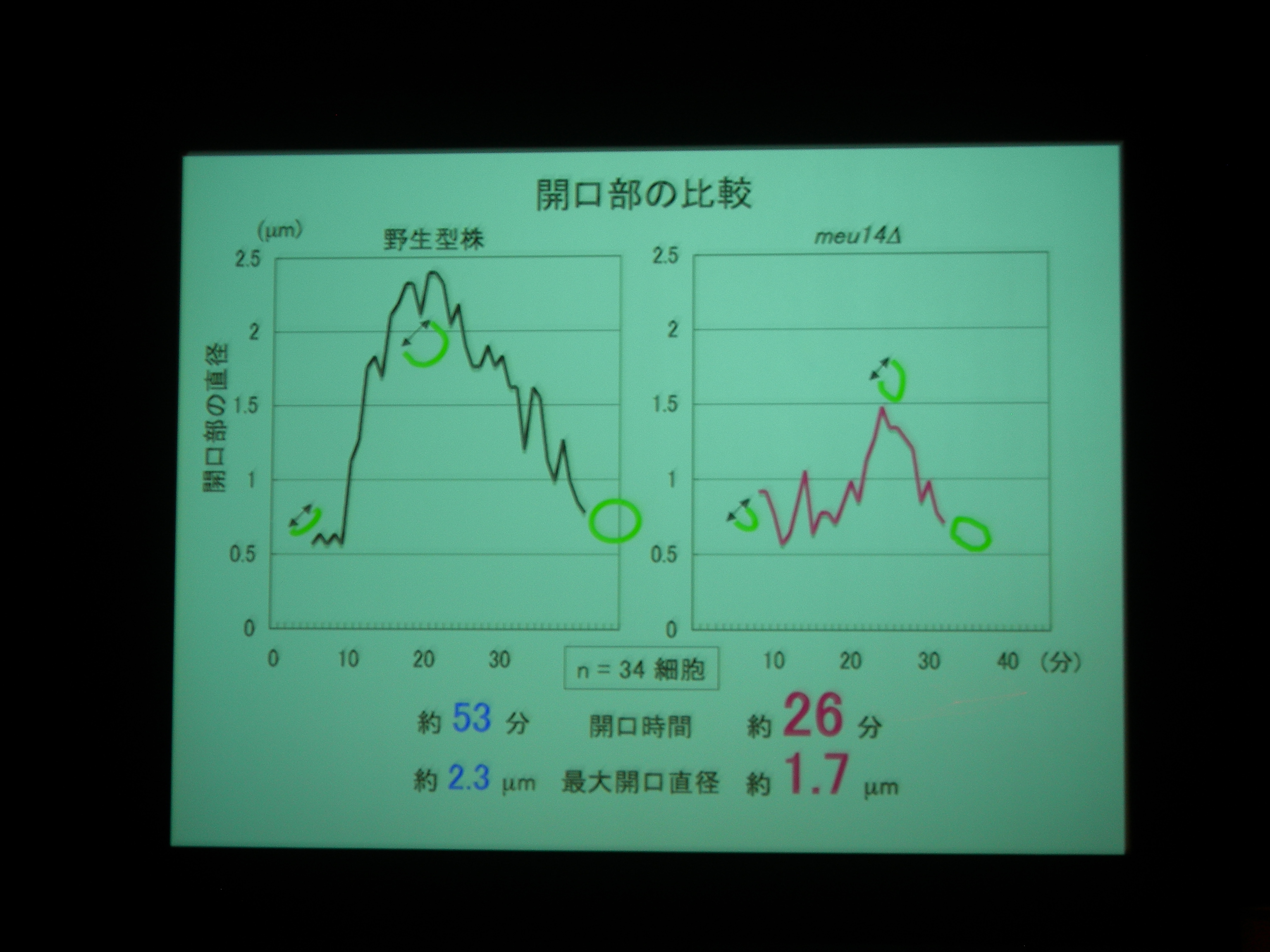
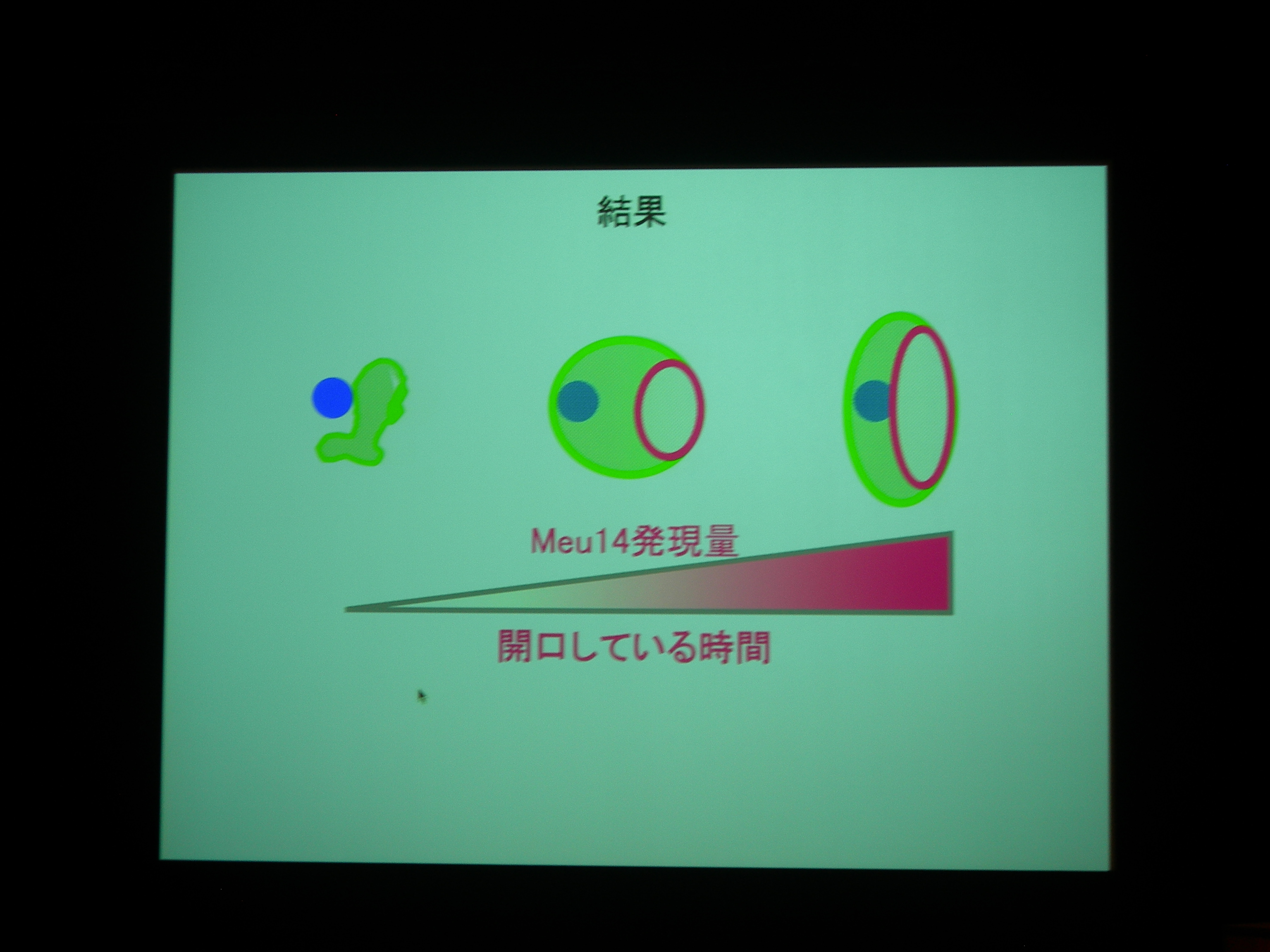
Meu14 を overexpression させると、口が大きく開き、開口している時間も長くなる (開くのにかかる時間は同じで、閉じるのに時間がかかる) が、まあ、わりとうまく動く。
開口は Meu14 の重合で、閉じるのは脱重合とすると、重合は濃度に依存し、脱重合は依存しないので、この現象が説明できる。
また、Meu14 は開口部をもたない膜には局在しないことが確認された。
脱重合のトリガになるような因子はなにかある? → まだわかっていません。
膜の開口部がどういう構造になっているかわかる?一重膜の開口部って? → あ、二重膜です。
動画を見ていると開口部どうしがおたがいにくっついて同調しているようにみえるんだけど → 相互作用しているかもしれないんだけど、近接している時間は非常に短いので、そうじゃないかも。離れていても同調して閉じます。
中身の核やなにかとの相互作用はないんですか?中身がちゃんとしたものであることを確認してから閉じているとかはない? → Meu14 がないと、核を分配するスピンドルが折れちゃったりするので、何かあると思う。
タグ: Biology
Synthesized Mycoplasma genitalium genome
Mycoplasma Genitaliumのゲノム合成に成功という記事がScienceに出てた。すごいな。
でも合成生物学もここまでくると、ちょっと怖い。