今日はちょっと時間をとって、自転車を整備することにした。何かと忙しくて後回しにしがちだけど、ちゃんとやらないと、あちこち傷むしね。
洗車して、予備のホイール (WH-R550) についていた古いタイヤをはがして、ハブをグリスアップしてリムフラップからタイヤまで交換。かなりの重労働だった。これでしばらく予備のホイールで走れるんだけど、後輪のフリー側はかなりグリスがどろどろになっており、ベアリングの球もちょっと曇っていたりして、心配な感じ。まあ、ボールレースは錆びてなかったし、大丈夫でしょうけれど。タイヤはずっと前に買ってあった Panaracer Extreme Evo2 と、Valiant Evo2. 同じのを2本買ってあるつもりだったんだけど、違ったか。ま、基本的に同じようなタイヤなので、予備としてはこれでいいでしょう。
ハブの調整は汗をかかない時期にやったほうがいい気がするので、気合い入っているほうのホイールも近々ちゃんとやろうと思う。が、今日は時間と体力がなくなったので、これでおしまい。
月: 2010年2月
昨日の自転車
雨降ったり体調崩したり出張したりして、3週間ぶりに乗った。
まだ胃の具合があんまりよくなくて、帰りは全然飛ばせない感じで、まあ、のんびり。
23.15km @ 23.6km/h (58m51s) odo 8041.4km
Decoding Read Completion Boundary register of PCIe
It’s important to know the RCB, read completion boundary to decode the completion TLPs for read request.
The RCB configuration is written in the Link Control Register, bit 3. The bit is 1’b0 if the RCB of root complex is 64 bytes, and 1’b1 for 128 bytes.
FYI, the bit appears on cfg_lcommand[3] on Xilinx’s Endpoint Block Plus.
PCIe
土曜の夕方からずっと胃腸炎で寝込んでいたわけですが、その間にも夢に PCI Express の transaction layer interface (Xilinx のやつ) の波形が出てきたりして、だいぶそっち方面でも病的だった。
昨日は死んだように眠っていたので、ほとんど何もしなかったんだけど、今日は布団の中で上半身起こして、わりとごりごりやって、RTL の波形ではかなりいい感じになった。少なくとも、Handshake はちゃんとできている感じ。
あとは、ヘッダをちゃんと作るとか、上位のモジュールとやりとりするための信号をいくつか作り込めばあとは、いま動いている PIO なインタフェイスとくっつける作業かな..
M-x verilog-mode
The comment out by C-c C-c is very cool:
/* -----\/----- EXCLUDED -----\/----- #(2*STEP) trn_tdst_rdy_n <= 1; #(1*STEP) trn_tdst_rdy_n <= 0; -----/\----- EXCLUDED -----/\----- */
Decoding TLP max payload size of Xilinx PCIe Core
While I’m writing a DMAC on my Virtex-5 PCIe card, I’ve found that the “MAX_PAYLOAD_SIZE” register has only 3 bits, while the size will be between 128 and 4096. And here’s the answer:
3’b000 128B
3’b001 256B
3’b010 512B
3’b011 1024B
3’b100 2048B
3’b101 4096B
3’b110 Reserved
3’b111 Reserved
(from Xilinx application note XAPP1052 documentation).
and now I can continue!
金沢→京都
金沢ではちょっと時間があったので駅前のスターバックスに寄って、福井行きに乗る。
外はすごい雪。

駅のホームは除雪が必要な状態。

車内は暖房が効いて、うたた寝するのが最高に気持ちいい。

福井まで乗ってきた電車は寝台電車を改造したやつ。もともと車内には3段ベッドがあったわけで、だからちょっと背が高くて、間延びした顔がかわいい。

そして、番号が1番でした。でも、この電車もそろそろなくなるとかそんな噂も。

福井では、福井新聞社ビルの1階にある「さくら通りカフェ」へ。駅前からはとりあえず路面電車に乗ってみた。

裁判所前で降りて、さくら通りをちょっと歩けばすぐ。花の季節はまだまだ遠いなあ。

ここのカフェは、オフィス街にあるわりとふつうのカフェなんだけど、テーブルの配置がとても広々しており、気持ちいい。普通なら2倍くらいテーブル置いちゃうでしょう、というくらいの密度。
ランチ前だったので、コーヒー一杯でずいぶん長い間仕事させていただきました。そうそう、MacBook の予備バッテリーまで持参してデータ処理しており、遊んでるわけじゃないのです。


帰りは福井城趾のほうを回って駅へ。
と、廃車.com って!! すごいな。

福井城趾は県庁か何かになっており、石垣の上に鉄筋の建物が建っている。かなりシュール。

敦賀行きの列車に乗ると、また降ってきた。しかもかなりの勢い。

敦賀の手前は北陸トンネル。すごく長い。
そういえばさっきと同じ型の電車だったので、車内のスナップを撮っておいた。
網棚の上にはベッドがあるわけだが、これ、中はどうなってるのかな。重たそうだから、いっそ外しちゃえばいいのにね。窓は、開くところと、普通の特急電車式にはめ殺しになっているところがあって、開くところに座ったら、すきま風が寒かった。



敦賀に到着。

おおっ、これが世界的に有名なオバマ線ですね。

近江塩津で乗り換えて、一路京都へ。長かった。

科研費ゲノム特定成果公開シンポ: Day 2
[ DNA はどのように折りたたまれているのか? ]
森下先生@東大。
源氏物語: 原本は存在しなくて、写本がたくさんあり、それぞれ巻数が違ったり indel とかがある → ゲノムの染色体の本数とか配列の違いに似ている?
ヒトのゲノムは 10 時間で複製される。50塩基/秒で、100並列。
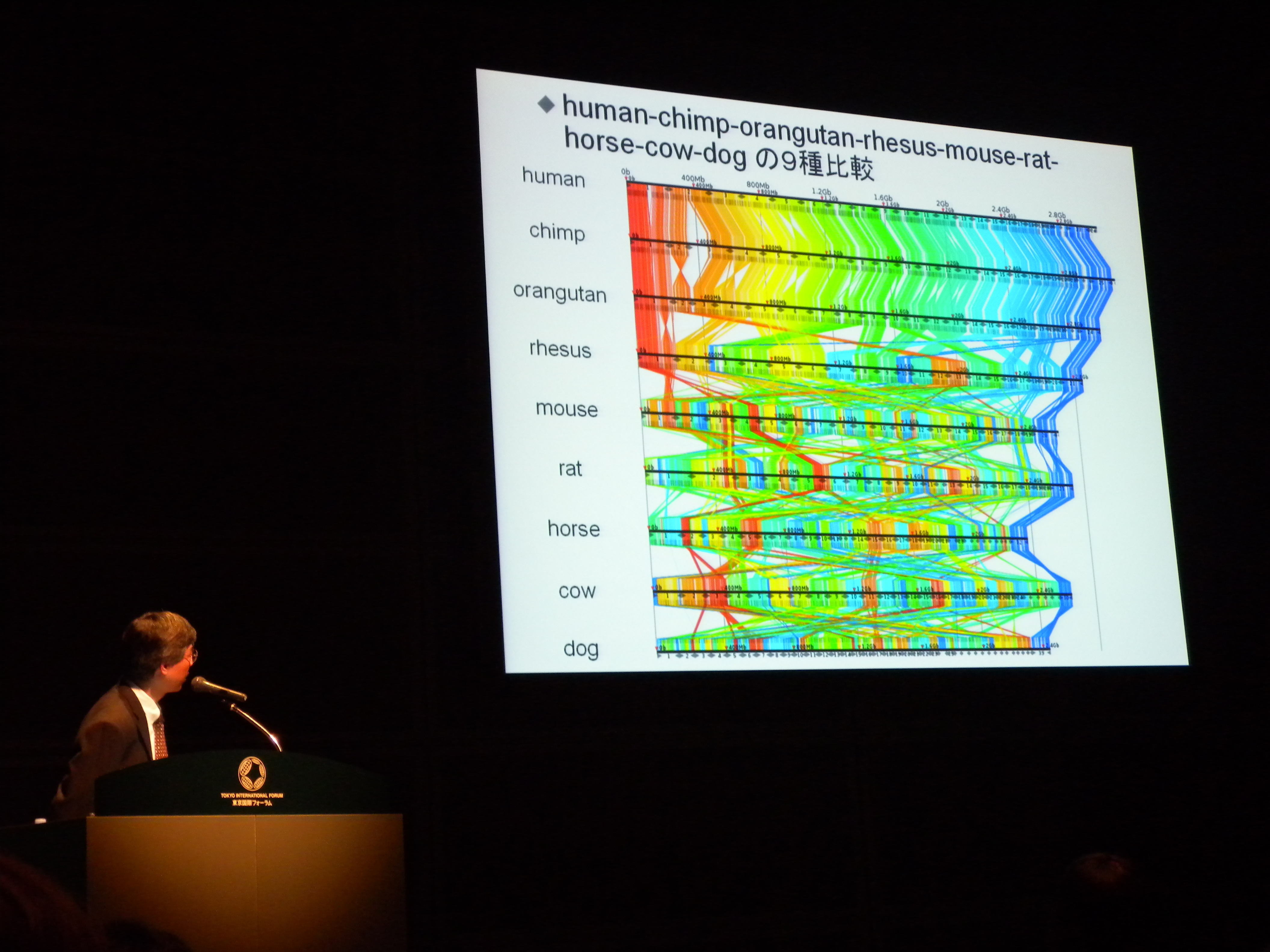
祖先ゲノムがどうだったか、ということを知るのに、ヒト・チンパンジーとホヤ・ナメクジウオだけではなくて真ん中の魚類くらいのが必要だった。
脊椎動物の祖先ゲノムは10本くらいの染色体があったのではないかと推定しており、ヒトに至るには2度のゲノム重複が起きている。魚類はさらにもう一度。
染色体が物語の一巻だとしたら、ページに対応するのがヌクレオソーム構造。
プロモーター結合がページを開くことに、メチル化がページを閉じることに例えられる。
(染色体をコピーするときはヒストンに巻きついた状態で複製され、体細胞の分裂の際にはメチル化の情報も伝わっているっぽい、のだそうです。)
メダカ+Solexa で調べてみた結果、ヌクレオソーム構造を取っているところとそうでないところで、塩基置換率が違う (ヌクレオソーム構造を取っているところでは、ヌクレオソーム構造を保存する方向に塩基置換が起きている。10年くらい前の酵母の研究では、ヌクレオソーム構造の中心ではリンカーの部分に比べて修復率が落ちているので、そっちが問題なのかも) ことがわかった。おもしろいな。
コンピュータはだいたい 2x in 18month なのが、sequencer は 2x in 8month なのでやばいね、というお話も。そうなんです…
Solexa は 1bp/h くらいで超並列だけど、Pacific bio のは 4bp/s くらい。ただ、後者は並列化が課題みたい。
中国はすごい勢いで Solexa GA とかを購入して頑張っているそうだ (台数としては、日本全体とあんまりかわらないけど、日本は分散しているのが問題かな。)。うおー。がんばらないとなあ。
[ Wiki によるデータベースと研究成果の発信 ]
有田先生@東大。
計測と解析は速くなった。でもその真ん中にある整理 (curation) は?
科学は知識の積み上げだから、成果が再利用されないと意味がない。でも、論文も学会発表も報道発表も、再利用には向いていない。
我々はデータ中心という新しいパラダイムに直面している。一般人は科学を信じ、期待しているのに、科学者は一般人に理解してもらうことを放棄。科学者がデータ中心の概念に対応できていないのは問題で、いつまで雑誌崇拝がつづくのか?
subscription fee が上がっている背景には、投資ファンドが雑誌の出資元を持っていることも影響している。
有田先生 Wiki はすでにいくつかの学会の公式なデータベースとして認定していただいているそうで、成果公開というのは大変重要なプロセスにおけるこういう活動は非常に重要だと思う。
数式とか理論的なものはいいけど、ゲノムとかそういう大規模なデータはどうやったらいいと思いますか (森下先生) → Wiki は backend が RDB なので、なにかそういうものを考え直さないといけない。いずれにしても Web ベースでやれる仕掛けは必要。
[ 多生物種のゲノムを高速に比較する並列システムの開発 ]
榊原先生@慶應大。
質疑だけメモ。
– マウス・ラットでは再編成が起こりやすい系統があるようですがそういうのと比べてみると面白いかもしれません。
– X染色体はヒトとチンパンジーでかなり保存されているがY染色体はシャッフルされている、という話がでているけれど、どうでしょう。 → Y のほうはまだアセンブリがきれいに進んでいない生物が多いので比較ができないのが現状です。
– NIG とか UCSC で visualize できなくて Murasaki & GMV でできるものは? 入力フォーマットは一般的なもの? → お答えしておきました。まさか回答させられるとは・・・
科研費ゲノム特定成果公開シンポ: Day 1
午後から参加。微妙に遅刻したところを U 先生に見つかってしまいました。えへへ。
廊下で立ち話して、最近の進捗をお伺いする。お手伝いできることがありそうなのでがんばります。
[ 土壌での微生物の生き様をゲノムから見る ]
津田先生@東北大。
土壌汚染物質を添加して、時系列でメタゲノム。菌叢が大きく変わるの、面白いな−。
しかし、メタゲノムやると、16s rRNA は 99% データベースに当たらない暗黒な世界なのだそうで、それはどうやったら解決できるのかね?
[ 麹菌のゲノム情報を活用した有用タンパク質の高生産 ]
五味先生@東北大。
「もやしもん」で微生物に興味を持つ高校生が多いのかー。
洗剤のアルカリ・リパーゼは麹菌の組み替え株で大量生産しているのだそうな。
固体培養 (麹培養) と液体培養では全然生産性が違う。低水分ストレスとかが効いてるのかな。
いろんな転写因子を網羅的に高発現させてみた。
麹菌に遺伝子導入をして工業的に使う、というのは既にいろいろやられているのだけれど、麹菌はプロテアーゼとかペプチダーゼを大量に作る (しかもものすごい種類を持っている!) ので、遺伝子導入してもせっかく作ったタンパク質を壊してしまう。
タンパク質分解酵素に関連する転写因子をみつけてそれを破壊してみたところ、多くのプロテアーゼの活性を抑えることができ、収量が改善した。
[ 病原性大腸菌の比較ゲノム解析とその応用 ]
林先生。
O157 ではふたつの IS (IS629, ISEc8) が飛び回っており、ファージのところに頻繁に入っている。
2回のPCRでO157であるかと、どの株であるかを特定することができるプライマーセットを作った。これは、集団感染であるかどうかを診断するために非常に有用で、制限酵素+パルスフィールドよりずっと速く、簡単。TOYOBOでキット化しており、診断に使われている。
日本では O157 >> O26 > O111 だけど Non-O157 の割合が増えてきている。欧州などではO157 の割合は比較的少なく O121 > O91 > O103 > O128 > …
O26, O111, O103 の配列を決定したが、ゲノムワイド系統解析では O157 とはまったく別になるものの、遺伝子クラスタに基づく解析だと EHEC は同じグループになり、並行進化していることがわかった。
Non-O157 EHEC のラムダ型ファージも、O157同様に III 型分泌器をもっていたり、プラスミドが同じような病原遺伝子セットをコードしている。でも、プラスミド間で相同性があるのは病原遺伝子の領域だけで、もともと同じプラスミドを獲得してきたわけではなさそう。
O157 とウシとの関わりはこれからやらないといけない。直腸の出口あたりにかなりの長期間定住しており、ウシでは病気は引き起こさない。一種の常在細菌といえるかも。
[ メダカの研究から発生、進化、病気のメカニズムを理解する ]
武田先生@東大。
メダカの内臓逆位個体では繊毛のモータータンパク質の欠失による腎臓病 (多発性嚢胞腎) や精子の運動性低下が起きる。ヒトでも繊毛の運動性低下が原因の遺伝性疾患があるが、同じ遺伝子の塩基多型が原因で発生することがわかった。
背腹の形成に異常がある変異体 (お腹も背中も丸くて銀色で、背びれと腹びれが同じ大きさ) もある。これの原因領域には2つの遺伝子があるだけだが、これらのコード領域は同じ。しかし、変異体では転写制御領域に変異があって発言パターンが異常になっていた。
メダカはいろいろな変異体が愛好家によって飼われていて、ゲノムも一倍体なのでいい感じ。金魚は四倍体?
[ 立襟鞭毛虫のゲノム情報から動物の多細胞化を探る ]
岩部先生@京大。
Monosiga ovata. 淡水性。64Mbp. 2万遺伝子。
海水性のは 42Mbp で、9000遺伝子くらい。
チロシンキナーゼは動物特異的だと思われてきたが、立襟鞭毛虫でも見つかっており、分子進化系統樹を書くと面白い。ほかにも動物特異的に多様化したと考えられていたシグナル伝達分子があり、それらは今後の課題。
イソギンチャク (刺胞動物): 神経系あり。
センモウヒラムシ (板状動物): 細胞が4〜5種類。神経系がない。
とかの微妙なところを含めて比較ゲノム解析してみた。
これすごいなー。
カドヘリンは僕らは細胞接着に使うけど、立襟鞭毛虫も持っている。襟のあたりでエサを捕まえるとか、なにか違うことに使っているのかも?
[ マウス亜種間ゲノム分化を利用した複合形質の遺伝解剖 ]
城石先生@遺伝研。
Mus musculus は地域ごとにいろんな亜種がある (北米には元々いなかった!)。
世の中で使われている実験系統 (C57BL/6J) は西ヨーロッパ産亜種。
遺伝研では三島でつかまえたマウスから日本産亜種の系統 (MSM/Ms: MSM=MiShiMa) を確立。 亜種間の塩基置換率は 0.87% くらい。
MSM/Ms はキャピラリーで 2.4x, Solexa で 15x で読んで貼り付けて、99% coverage. 11,742,976 SNPs. うち、Non-synon. SNPs (アミノ産置換あり) が 26.000. Non-sense mutations は 131 遺伝子で見つかった (0.5%!)。
MSM/Ms は体のサイズが約半分、発がん抵抗性が高い、エネルギー代謝系が節約型、など、表現形のレベルでいろいろな違いがある。
亜種間交配によって特定の染色体だけが別の亜種由来なもの (コンソミック系統) をすべての染色体について作った。
多くの量的形質については、単に足し算と引き算だけで成り立つ (全部足したら100%になる) のではなく、全部足すと100よりずっと大きくなる (遺伝的交互作用=epistatsis が強い) ことがわかった。
金沢行き
試験監督が終わってから新宿のニコンに寄って帰宅して、慌ただしく準備して23時に上野駅。13番ホームには金沢行き寝台特急「北陸」が入線しており、写真を撮る人たちであふれかえっている。23時3分発。


夜行列車の廃止は、この何年かでじわじわ進んできて、いよいよ全滅に近い感じなのだけれど、馴染みの土地への列車がなくなるというのは切ない。夜、上野や東京や新宿といったターミナルへ行けば、いつでもそこへ行ける、という、精神的な保険みたいなものがなくなる気分だ。僕にとっては、中央線の急行「アルプス」の廃止がそれだった。故郷へつながっている細い糸を断ち切られる、そんな感覚。
僕は同じ金沢行きの、23時33分発急行「能登」に乗る。こちらは座席で、どちらも3月のダイヤ改正で消える。時刻表の「3月12日まで運転」の文字がせつない。


昔ながらの寝台車に乗れる機会ももうあとわずかなのだなあ、と思いつつ、最後に「能登」を選んだのは、学生時代からの貧乏性が抜けないというか、それが自分の旅のスタイルとして染みついているからなのか。
いつもは高崎くらいまで起きているのだけれど、あまりの疲労に耐えきれず、熊谷の手前でボックスシートに、カバンを枕に L 字になって気絶。急行列車とはいえ、車両は往年の特急型であり、暖房がしっかり効いて非常に快適。ひと昔前みたいに、冬は窓のすきま風で寒くて眠れない、なんていうこともない。
途中で一度目を覚ましたときは、上越国境のトンネルの中。本当にトンネルを抜けると雪国であり、夜の底が白くなった。スキー場を眺めて眠りに落ち、再び目が覚めるとまだ糸魚川で、ダイヤ通りならもう富山に着く頃合いだが、新潟県内が大雪で1時間くらい遅れている、というアナウンス。外は雪景色。

朝目が覚めると雪景色で、知らない土地を走っている、という旅ができなくなる日も、そう遠くない気がする。時代の流れといえばそれまでなのかもしれないけど、残念だ。
新幹線や飛行機でびゅーんと行ってしまうには、この国はあまりにも美しすぎる。
あと1ヶ月。
もう一度くらい乗れるかな。
